Difference between revisions of "Davidson/Missouri Western iGEM2008"
MaCampbell (talk | contribs) |
MaCampbell (talk | contribs) |
||
| Line 9: | Line 9: | ||
==[[Contact A Team Member]]== | ==[[Contact A Team Member]]== | ||
| − | == | + | == [[TeamProject]] == |
==[[Wet Lab Pages]]== | ==[[Wet Lab Pages]]== | ||
Revision as of 19:49, 13 June 2008
Davidson College - Missouri Western State University
iGEM 2008
Contents
- 1 Contact A Team Member
- 2 TeamProject
- 3 Wet Lab Pages
- 4 Math Modeling Pages
- 5 Las/Rhl cell signaling system
- 6 Lux cell signaling system
- 7 The ainS Quorum Sensing System???
- 8 Lsr (AI-2) cell signaling system
- 9 Fec cell signaling system
- 10 Signal molecules
- 11 E. coli Signaling
- 12 Cell signaling resources
- 13 Davidson Journal Club
- 14 iGEM 2007 Useful Information
- 15 iGEM 2006 Useful Information
- 16 iGEM 2005 Useful Information
Contact A Team Member
TeamProject
Wet Lab Pages
Math Modeling Pages
Las/Rhl cell signaling system
Responsible: Robert Cool, Alicia Allen, and Erin Feeney
Las System
Signal Molecule: An AHL called PAI-1 (N-3-oxododecanoyl-l-hsl)(3-oxo-C12-hsl)
Bacterial species: Pseudomonas aeruginosa gram(-) possibly E.coli (see article 3)
Receiver protein: LasR
Effect of binding: TXN activation of virulence genes, lasA, lasB, apr, toxR
Synthase: LasI enzyme
Target Genes: lasI, lasA, lasB, apr, toxR
Regulation: unknown
References
"Roles of Pseudomonas aeruginosa las and rhl quorum-sensing systems in control of elastase and rhamnolipid biosynthesis genes"
JP Pearson, EC Pesci and BH Iglewski [1]
"Regulation of Pseudomonas Quinolone Signal Synthesis in Pseudomonas aeruginosa"
Dana S. Wade, M. Worth Calfee, Edson R. Rocha, Elizabeth A. Ling, Elana Engstrom, James P. Coleman, and Everett C. Pesci 2
Posttranscriptional Control of Quorum-Sensing-Dependent Virulence Genes by DksA in Pseudomonas aeruginosa
Florence Jude,Thilo Köhler,Pavel Branny,Karl Perron,Matthias P. Mayer,Rachel Comte, and Christian van Delden 3
Pending: [2]
Rhl System
Signal Molecule: An AHL called PAI-2, Plasminogen activator inhibitor-2, N-butanoyl-homoserine lactone (C4HSL)
Bacterial species: Pseudomonas aeruginosa, gram(-)
Receiver Protein: Rhl R
Effect of Binding: activation of Rhamnosyl Transferase, then making RL (rhamnolipid)
Synthase: RhlA and RhlB
Target Genes: pqsABCDE and phnAB
Regulation: unknown
References
background information on Las and Rhl

Parts Needed:
LasR + pro/term
RhlR + pro/term
RhlI + pro/term
pqsABCDE + pro/term
pqsR
pqsH + pro/term
phnAB
LasI
RBS
Lux cell signaling system
Responsible: Andrew Gordon and Pallavi Penumetcha
Signal molecule: N-acyl-homoserine lactone (AHL) Generic term for a variety of species specific hormone-like molecules
Bacterial species: discovered in Vibrio fischeri known to work in E. coli
Receiver protein: LuxR protein receives signal from AHL; also has some control over transciption of luciferase
Signal molecule synthase: LuxI; also has some control over transciption of luciferase
Additional Information: "Quorum Quenching" aiiA (intracellular) lactonase reduces AHL concentration

Resources
AHL signaling molecules by species; some are specific to gram pos but may affect gram negs
References
Quorum Quenching to control Lux Pathway
[3] A Synthetic multicellular system for programmed pattern formation
Parts Needed:
LuxR + pro/term
RBS
LuxI + pro/term
LuxI sender
The ainS Quorum Sensing System???
References
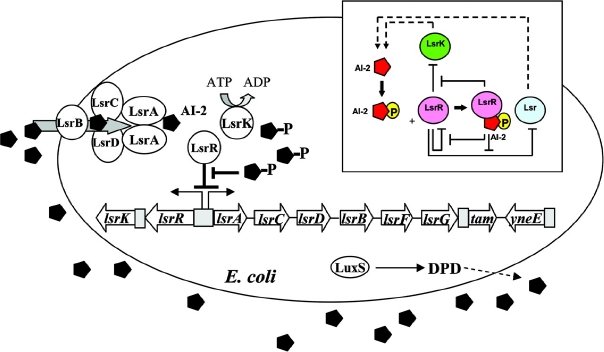
Lsr (AI-2) cell signaling system
Responsible: Kelly Davis, Xiao Zhu
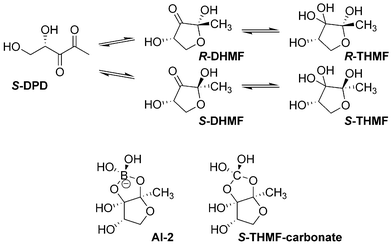
Signal molecule: AI-2 (furanosyl borate diester in V. harveyi, a variety of other molecules in other species), all are derived from DPD [4]
Bacterial species:
lsrA,B,C,D,F,G,R,K: Escherichia coli HS, SMS-3-5, str. K12 substr. MG1655, and substr. DH10B.
lsrE:Escherichia coli str. K12 substr. MG1655
LuxS:Escherichia coli HS, SMS-3-5, APEC O1, str. K12 substr. MG1655, substr. DH10B, and UTI89.
Receiver protein: LsrR protein receives signal from sensor protein
Signal molecule synthase: Pfs enzyme, then LuxS autoinducer synthase
Target genes: lsr operon, including ABC transporter and LsrK kinase
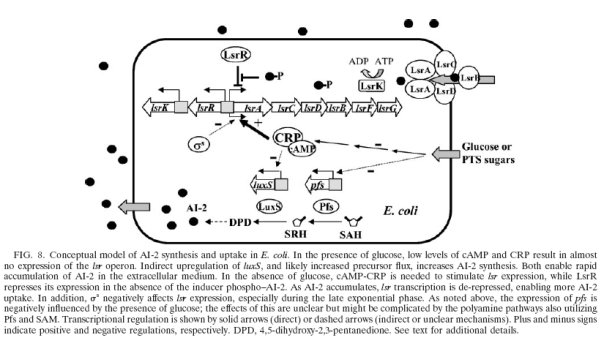
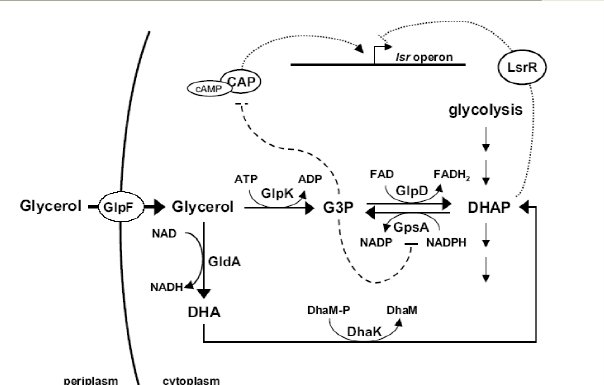
Regulation: LsrR represses the lsr operon, derepression by phospho-AI-2; catabolite repression influences AI-2 accumulation through the cyclic AMP (cAMP)-cAMP receptor protein (CRP) complex, which directly stimulates transcription of the lsr operon and indirectly represses luxS expression.cAMP-CRP is shown to bind to a cAMP receptor protein (CRP) binding site located in the upstream region of the lsr promoter and works with the LsrR repressor to regulate AI-2 uptake.
Note: AI-2 is synthesized and secreted during exponential growth and is imported in stationary phase when glucose becomes limiting. In the presence of glucose, AI-2 is not imported because the lsr operon is not transcribed due to camp-CAP mediated repression. Both glycerol and G3P(glycerol 3-phosphate) repress lsr transcription, while the majority repression comes from G3P. DHAP represses lsr transcription by a cAMP-CAP-independent mechanism involving LsrR.
Parts Needed:
LsrR pro/term
LsrK
LsrACDB (transport)
LsrFGE (catabolic)
LuxS
Pfs enzyme (?)
http://gcat.davidson.edu/GcatWiki/images/b/b8/N654260305_1291548_2335.jpg
{kind=link}
Note:
lsrB encodes the periplasmic AI-2 binding protein
lsrC & lsrD encode the channel proteins
lsrA encodes the ATPase that provides energy for AI-2 transport
lsrF is similar to genes specifying aldolases
lsrG encodes a protein with an unknown function.
tam: trans-aconitate 2-methyltransferase, also known as lsrE or yneD yneE:conserved inner membrane protein
http://gcat.davidson.edu/GcatWiki/images/3/30/N654260305_1340817_550.jpg
{kind=link}
 R-THMF
http://gcat.davidson.edu/GcatWiki/images/8/8c/S-DPD.gif
http://www.nature.com/nrmicro/journal/v3/n5/images/nrmicro1146-f2.gif
R-THMF
http://gcat.davidson.edu/GcatWiki/images/8/8c/S-DPD.gif
http://www.nature.com/nrmicro/journal/v3/n5/images/nrmicro1146-f2.gif
{kind=link}
{kind=link}
http://gcat.davidson.edu/GcatWiki/images/c/c9/Grl.jpg
{kind=link}
DHAP: dihydroxyacetone phosphate.
http://gcat.davidson.edu/GcatWiki/images/9/9b/Grlw.jpg
{kind=link}
References
Signaling explained with graphics of AI-2 pathways
Lsr-mediated transport and processing of AI-2 in Salmonella typhimurium
Review of AI-2 and other systems
Regulation of Uptake and Processing of the Quorum-Sensing Autoinducer AI-2 in Escherichia coli
Resources
lsr nucleotide sequence in EcoCyc
Transcription of LuxS (interaction with micA,gshA)
http://gcat.davidson.edu/GcatWiki/images/c/cb/N654260305_1347432_6212.jpg
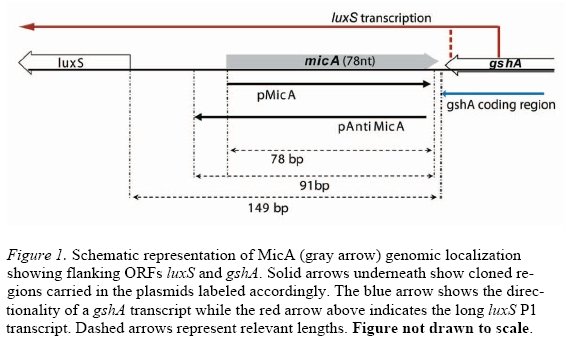{kind=link}
Fec cell signaling system
Responsible: Xiao Zhu, James Barron (DC)
Ferric Dicitrate Transport System
The inducer, ferric citrate, binds to an outer membrane transport protein, FecA, and without further transport elicits a signal that is transmitted across the outer membrane (by FecA), the periplasm, and the cytoplasmic membrane (by FecBCDE and FecR) into the cytoplasm. Signal transfer across the three subcellular compartments is mediated by the outer membrane transport protein (FecA) that interacts in the periplasm with a cytoplasmic transmembrane protein (FecR). FecR is required for activation of a sigma factor (FecI) which belongs to the extracytoplasmic function (ECF)sigma factor family.
Only iron not iron complex enters the cytoplasm. FecA is the TonB energy transducing system-dependent.
Signaling Molecule: FeC (ferric dicitrate)
Bacteria species: E.coli, Pseudomonas putida, P. aeruginosa, Serratia marcescens, Klebsiella pneumoniae, Aerobacter aerogenes, Bordetella pertussis, B. bronchseptica, B. avium, and Ralstonia solanacearum.
Receptor Protein: FecA
Effect of binding: the conformational changes that FecA undergoes when binding to ferric citrate:The alpha helix in loop 7 unravels, and the loop moves by up to 11 angstroms ; Loop 8 moves up to 15 angstroms.
http://www.rsc.org/ej/CS/2007/b617040b/b617040b-f13.gif
Sensor Producer: N/A
{kind=link}
Harvard iGEM'07 team worked with Fec system, the results were not favorable. :We believe that overexpression of the Fec system killed the cells, possibly by disturbing the cell membranes.
Regulation of the FecI-type ECF sigma factor by transmembrane signaling
[5]
More detailed information about Fec
Exogenous Induction of the Iron Dicitrate Transport System of Escherichia coli K-12
Control of the Ferric Citrate Transport System of Escherichia coli:Mutations in Region2.1 of the FecI ECF Sigma Factor Suppress Mutations in the FecR Transmembrane Regulatory Protein
Docking of the Periplasmic FecB Binding Protein to the FecCD Transmembrane Proteins in the Ferric Citrate Transport System of Escherichia coli
Interactions between the Outer Membrane Ferric Citrate Transporter FecA and TonB: Studies of the FecA TonB Box
Signal molecules

Gram (-) bacteria use:
3-oxo-C6-HSL, N-(3-oxohexanoyl)-L-homoserine lactone, an AHL
DPD, the AI-2 precursor, 4,5 dihydroxy-2,3-pentanedione
HHQ, 2-heptyl-4(1H)-quinolone, an AQ
Gram (+) bacteria use:
DPD, the AI-2 precursor, 4,5 dihydroxy-2,3-pentanedione
A-Factor, 2-isocapryloyl-3-hydroxymethyl--butyrolactone
PQS, pseudomonas quinolone signal, 2-heptyl-3-hydroxy-4(1H)-quinolone
DSF, ‘diffusible factor’, cis-11-methyl-2-dodecenoic acid
3OH-PAME, hydroxyl-palmitic acid methyl ester;
AIP-1, staphylococcal autoinducing peptide 1
E. coli Signaling
"As yet no AHL-producing Escherichia coli or Salmonella strains have been identified, although both organisms possess an AHL receptor (SdiA) of the LuxR protein class and respond to AHLs produced by other bacteria." Williams 2007
Cell signaling resources
Featured parts in iGEM registry
Quorum sensing, communication and cross-kingdom signalling in the bacterial world
Bonnie Bassler lab at Princeton
One-Stop Shopping for QS from Nottingham
Special Issue: Bacterial conversations: talking, listening and eavesdropping
Dialogs with Bacteria: Quorum Sensing
General Types of Cell Signaling: Bacteria on Steroids?
Modeling Cell-Signaling Networks
Quorum Sensing and the Population Control of Virulence
Davidson Journal Club
Stochasticity and Gene Expression --- Dr. Campbell
iGEM 2007 Useful Information
Virginia Tech
Engineering and Epidemic
The use of bacteria to model the spread of a disease. It would appear that cell-to-cell communication is a major part of the design of the project. It is unclear how successful the team was in building parts useful to us. Most of the project seems to be on the mathematical modeling side of things.
The use of bacteria to model the spread of a disease. It would appear that cell-to-cell communication is a major part of the design of the project. It is unclear how successful the team was in building parts useful to us. Most of the project seems to be on the mathematical modeling side of things.
http://parts.mit.edu/igem07/index.php/Virginia_Tech
University of Waterloo
Half-Adder Logic Gate
The goal of this project is to design a basic device for computing. Our idea was to reproduce a circuit element called a half adder with DNA, which takes in two 1-bit inputs, adds them, and outputs a sum and a carry. Our device responds to two inputs: red light and the chemical tetracycline. The input sensors control a set of genetic switches in order to carry out the computation and fluoresces green, red, or neither, depending on the outcome. Useful for long addition in base-2.
http://parts.mit.edu/igem07/index.php/Waterloo
UCSF
Project 1: Protein Scaffolds as a Molecular Breadboard
Using synthetic protein scaffolds to control information flow of a kinase pathway in eukaryotic cells.
http://parts.mit.edu/igem07/index.php/UCSF
Tianjin
Biological diode
In this project, we try to construct a biological device to imitate the function of the diode, one of the most significant parts in the electric integrate circuit. The flow of molecular signal AHL is considered as the current of electric circuit. The generator, amplifiers, blocks and detector cells are constructed with the parts provided by MIT and then are equipped in series in order to establish the cellular and molecular biological diode. Our device, which is a combination of technologies from the field of computer science, molecular biology and chemical engineering, is a breakthrough for the application of mature techniques of chemical engineering to the field of synthetic biology.
http://parts.mit.edu/igem07/index.php/Tianjin
Duke University
Bacterial Communication With Light
http://parts.mit.edu/igem07/index.php/Duke/Projects/bc -
University of Cambridge
BOL: Bacteria OnLine
They talk a little about making a bacterial internet, I have no idea what they mean.
http://parts.mit.edu/igem07/index.php/Cambridge
Tokyo Tech
Pareto's Principle: An Ant Society
The goal of our project is to make a bacterial society that follows Pareto's principle as an ant society does. On the other word, we try to construct a bacterial system which takes "balanced differentiation". Bistability and cell-cell communication are necessary to realize our model of "Balanced differentiation".
http://parts.mit.edu/igem07/index.php/Tokyo_Tech
Quorum Sensing
See this quorum sensing web page
Harvard
Quorum Sensing
Was developing a luxL luxR quorum sensing system using OHHL. Lux quorum-sensing works like a system of sender and receiver.
http://parts.mit.edu/igem07/index.php/Harvard#Quorum_Sensing
Chiba
Communication Unit
Something about cell to cell communication involving LuxL, LuxR, and AHL. Hard to understand because they did not translate into English very well.
http://parts.mit.edu/igem07/index.php/Chiba/Communication
Brown
Cellular Lead Sensor
-no useful information
http://parts.mit.edu/igem07/index.php/Brown
Colombia-Israel (ORT Ebin High School)
A Microbial Biosensor Device
No description left...
- http://parts.mit.edu/igem07/index.php/Colombia-Israel%20(ORT%20Ebin%20High%20School)
Edinburgh
Division PoPper and Self Flavouring Yoghurt
- This team is working on a project that is looking into a form of cell communication
- "We designed a signal generator device that produces an output in the form of PoPS pulses each time a bacteria undergoes cell division. Therefore it may trigger actions as a function of cell replication."
- Could not find where on this page this info came from, but it was included with this link: - The goal of this project is to design a basic device for computing. Our idea was to reproduce a circuit element called a half adder with DNA, which takes in two 1-bit inputs, adds them, and outputs a sum and a carry. Our device responds to two inputs: red light and the chemical tetracycline. The input sensors control a set of genetic switches in order to carry out the computation and fluoresces green, red, or neither, depending on the outcome. Useful for long addition in base-2.
http://parts.mit.edu/igem07/index.php/Edinburgh#The_Projects.21
Imperial
Infector Detector
-no useful information, but really interesting project...
http://parts.mit.edu/igem07/index.php/Imperial + UCSF (2007)
Middle East Technical University
Chase simulator
This project was not completed, but has some interesting information on E. coli cells triggering a response in nearby E. coli cells. http://parts.mit.edu/igem07/index.php/Chase_Simulator
iGEM 2006 Useful Information
UT Austin 2005/2006 Project : Edge Detector Link to parts: http://parts.mit.edu/r/parts/partsdb/pgroup.cgi?pgroup=iGEM&group=iGEM_UTAustin
Useful information: They have "black boxed" the light-system and used it as an input for the of the edge detection circuitry.
Edge Detector Circuit and logic. The light sensing machinery from above has been black-boxed and the edge detection circuitry has been added downstream. Red light represses the expression of 2 genes; a biosynthetic gene for a membrane diffusible quorum sensing activator (AHL), and a dominant transcriptional repressor (cI). (Right) The output of the circuit (Z;Beta-galactosidase) is ON only in the presence of X (AHL) and the absence of Y (cI). This can only occur at the light/dark boundary.
Note: Built on 2005’s work. Pretty much the same as 2005.
Harvard “Cell Surface Targeting” http://parts.mit.edu/wiki/index.php/Harvard_2006
Project Overview “In order to target nanostructures to cells, we developed adaptamers, universal nucleic acid adaptars which can link two substrates. • Such an interface could also be used to link together entire cells for the study of cell-cell interactions and the linkage of two interacting proteins, in effect creating a nucleic acid enzyme. • Adaptamers generally depend on aptamers, short sequences of nucleic acid that bind with high specificity and affinity to particular substrates. • Tahiri-Alaoui et al. created the first aptamer in 2002, consisting of two aptamer sequences linked together by a bulky basepairing region ~100 nucleotides long. • Our goal was to create an adaptamer that could link together streptavidin and thrombin. Delivery of thrombin to a streptavidin-coated magnetic bead would show the potential for delivery of a macromolecule to a cell surface. Additionally, we wished to be able to be able to quench adaptamer function through the addition of an adapatamer-disabling oligonucleotide.”
The University of Calgary 2006 iGEM team is working on the following project. A petri plate is inhabited by two strains of genetically engineered E. coli bacteria. The first strain---the Senders---have been engineered to emit two chemical signals into the plate environment: Aspartate and Acyl Homoserine Lactone (AHSL). The senders themselves are activated by light. The second strain---the Receivers---have been designed to respond to each of these signals in a different way. The Receivers express Green Fluorescent Protein in the vicinity of AHSL. The Receivers also move towards areas of greater Aspartate concentration. The same bacteria also decrease Aspartate levels where they are present, as this is a nutrient and constitutes the reason for why they are attracted to it in the first place. Our goal is to make the Senders and Receivers create interesting behaviour dynamics visualized by fluorescent patterns.
http://parts.mit.edu/r/parts/partsdb/pgroup.cgi?pgroup=iGEM2006&group=iGEM2006_Calgary
Berkeley: networks of cells communicating via conjugation; demonstrated the transmission of a coded message
http://parts2.mit.edu/wiki/index.php/University_of_California_Berkeley_2006
“We have developed the process of addressable conjugation for communication within a network of E. coli bacteria. Here, bacteria send messages to one another via conjugation of plasmid DNAs, but the message is only meaningful to cells with a matching address sequence. In this way, the Watson Crick base-pairing of addressing sequences replaces the spatial connectivity present in neural systems. To construct this system, we have adapted natural conjugation systems as the communication device. Information contained in the transferred plasmids is only accessable by "unlocking" the message using RNA based 'keys'. The resulting addressable conjugation process is being adapted to construct a network of NAND logic gates in bacterial cultures.”
Mexico: cellular automata
http://parts2.mit.edu/wiki/index.php/IPN_UNAM_2006
“We wish contribute to the iGEM project development various protein based bio-components. We will work along three main lines: complex and reversible dynamical systems and formal languages, that support particles and multiple reactions, related to the molecular transformations.”
“We study two-dimensional cellular automaton, where every cell takes states 0 and 1 and updates its state depending on sum of states of its 8 closest neighbors as follows. Cell in state 0 takes state 1 if there are exactly two neighbors in state 1, otherwise the cell remains in state 0. Cell in state 1 remains in state 1 if there are exactly seven neighbors in state 1, otherwise the cell switches to state 0. CA governed by such cell-state transition rule exhibits reaction-diffusion like pattern dynamics, so we call this Diffusion Rule.”
“Using the diffusion rule we can generate a dynamical pattern over a system, like turn on/off ligth with alive o dead cells that shows a luminescence, examples include fluorescence, bioluminescence and phosphorescence.” “Starting with any configuration, the cells alive are represented in yellow (the activator) and dead in black (the inhibitor), see figure 4. The system is created defining an inicial state over the base configuration (see figure 3). The luminescence is obtained by the evolution of this initial pattern.”
Brown:Bacterial Freeze Tag http://parts.mit.edu/wiki/index.php/Brown:Bacterial_Freeze_Tag#Overview 2006 igem
This project involves programming bacteria to be able to play a game of freeze tag. Bacteria will be engineered to swim around a microfluidics device until they reach a certain proximity to the 'IT' cell and then they will lose their ability to move. This loss of motility will be combined with a change in color from Green to Blue. When another bacterium, which is moving (not the 'IT' cell), reaches a certain proximity to the 'frozen' bacteria it will again regain its ability to move and turn from Blue to Yellow.
TetR promoted with LuxI downstream. LuxI is an enzyme that produces AHL and will produce the red fluorescent protein (RFP). The AHL produced is exported from the cell where it then forms a complex with the LuxR protein that is produced by the AHL sensor within the Receiver cell.
The AHL sensor is TetR promoted and forms the LuxR protein which then forms a complex with AHL. This LuxR and AHL complex then activates the pLuxR promoter. Downstream of the pLuxR promoter is the LacI protein. LacI inhibits the pLac promoter on the "Freeze Machine".
A promoter that is regulated by LacI will promote the production of LasI, MotB, and cI. This will subsequently inhibit the production of CFP and LasR. In the presence of LacI, however, MotB, LasI, and cI will not be produced. CFP will therefore be produced along with LasR and LacI. This results in the "freezing" of the cell.
McGill University Split YFP
http://parts.mit.edu/wiki/index.php/McGill_University_2006
The idea behind the project is fluorescence complementation, which involves the joining of two leucine zipper proteins, Fos and Jun, each fused to a half terminus of YFP. Originally, the Fos and Jun proteins were fused to a beta gene coding for a membrane protein. The project involved performing a PCR reaction to produce two inserts, the N-terminus and the C-terminus of YFP, and then ligating these inserts into 2 vectors, containing Jun-beta and the Fos-beta respectively. The two fusion proteins (Fos-beta-YFPC and Jun-beta-YFPN) were expressed in the cell membrane of two populations of E. coli. We then allowed these two cell types to combine, resulting—ideally—in the complementary binding of the Jun and Fos proteins when the cells are in close contact. Consequently, the two half YFP fragments bind to form full YFP, and the cells will fluoresce.
Penn State
http://openwetware.org/wiki/IGEM:PennState/2006
The bacterial relay race takes advantage of an ability to control cellular motility using inducible promoters such as those involved in nutrient catabolism or quorum sensing. “Receiver” bacteria move in response to small-molecule signals either added to the system or originating from motile, “sender” strains. The most significant challenges relating to this project stem from difficulties of tightly controlling the target motility gene motB. Low levels of motB expression result in system failure (constitutive motility), and resolving this issue is essential to developing reliable modular systems that are the hallmark of synthetic biology
Tokyo http://parts.mit.edu/wiki/index.php/Tokyo_Alliance:_Conclusion
Our project is to make this Noughts-and-Crosses in vivo. -1. Inputs -1. Chemicals -1. To indicate each square -1. To be spreaded into all squares. -1. Outputs -1. Reporter of SYANAC: GFP Reporter of Human: RFP
We can say we will expand the number of regulator genes we can use to build logic gates and through this project we made simple constructing method.
BU 2006
Project: build a functioning "Biological Night-Light" system
Link to parts : http://parts.mit.edu/r/parts/partsdb/pgroup.cgi?pgroup=iGEM2006&group=iGEM2006_BU Goal Isolate luxCDABE and add the 4 BioBrick restriction sites to the ends of the gene. Ideas "Proteins that affect the wavelength of the emitted light, lumazine and yellow fluorescent protein, have been isolated from Photobacterium and Vibrio species, respectively. The lumazine proteins shift the color of the light to wavelengths shorter than 490 nm..." (Meighen 1991) Perhaps we could build a circuit to modulate the emitted wavelength by periodically expressing a carefully-chosen fluoresent protein. Think FRET and BRET.
Let's modify the lux operon so our bacteria can play Conway's Game of Life. In the game, discrete "cells" interact with one another according to four extremely simple rules, which essentially boil down to this: if a cell has too many or too few neighbors it turns off, otherwise it turns/stays on. These rules and the initial state of all the cells often produce systems of fascinating and lifelike complexity. Perhaps we could add a circuit such that LuxI would only be activated in response to a narrow "medium" range of concentrations of its autoinducer (3OC6HSL), not too much or too little. In fact, I think such a circuit has already been built by the Weiss lab and demonstrated with their infamous bullseye.
Weiss Lab: Game of Life Link: http://www.princeton.edu/~rweiss/ Note: Weiss Lab build a system that enables cells to “play” Conway’s Game of Life, where cells live or die based on the density of their neighbors. This system exhibits complex global emergent behavior that arises from the interaction of cells based on simple local rules.
Another system is a pulse generator where sender cells communicate to nearby receiver cells, which then respond with a transient burst of gene expression whose amplitude and duration depends on the distance from the senders. In another system, receiver cells have been engineered to respond to cell-cell communication signals from senders.
Bangalore NCBS 2006
Synchronization of bacterial cell cycles. Use a cell cycle-dependent promoter to drive a LuxI-LuxR based cell-cell signal. Use regulation of replication initiator DnaA to modulate cell cycle in receiver cells. Immediate goals: To determine if candidate promoters oscillate; to regulate DnaA levels
http://parts.mit.edu/wiki/index.php/Workshop
Rice University 2006 The objective of this project is to engineer Escherichia coli which are able to actively pursue and mark or eliminate another bacterial target. This system can be divided into three components: an input element, a processing element, and a response element. The input element will consist of a quorum sensing circuit which would allow specific detection of the bacterial target. The processing element will facilitate the signaling of this input into controlled responses. A number of different response elements can be conceived, to be used separately or in tandem: 1) integration into the chemotactic pathway of E. coli, allowing for directed mobilization towards the target, 2) reporter response at high pheromone concentrations to allow for visual identification of the target location (e.g., GFP production), and 3) an elimination response to produce molecules which are specifically lethal to the desired target. http://parts.mit.edu/wiki/index.php/PROJECT_PROPOSAL
Cambridge: http://parts.mit.edu/wiki/index.php/Cambridge_University_2006
The type 1 cell produces 3O-C6-HSL (represented by the small yellow cannon ball) while type 2 produces 3O-C12-HSL (represented by the blue cannon ball). The type 1 cell responds to 3O-C12 HSL and type 2 responds to 3O-C6 HSL. The response of type 1 cells can be visualized through the expression of RFP. The response of type 2 cells can be visualized through the expression of GFP.
1. Parts used for generating patterns (these are parts whose function Cambridge characterized) (a) Constitutively expressed fluorescent proteins: ECFP: BBa_I13601 GFP: BBa_J04430 EYFP: BBa_I6031 mRFP1: BBa_J04450 (b) Constitutive or auto-induced AHL synthesis: Lux-sender (auto-inducing): BBa_I15030 Las-sender (constitutive): BBa_I0407 Rhl-sender (constitutive): BBa_I0405 Cin-sender (constitutive): BBa_I0409 (c) AHL-induced fluorescence response: Lux-receiver (GFP): BBa_T9002 Lux-receiver (EYFP): BBa_I13263 Las-receiver (EYFP): BBa_I0426 Rhl-receiver (EYFP): BBa_I0424 Cin-receiver (EYFP): BBa_I0428
Princeton: http://parts.mit.edu/wiki/index.php/Princeton:Project_Summary
Mammalian cell-cell signaling using LuxR and LuxI…not applicable
iGEM 2005 Useful Information
Caltech http://www.cds.caltech.edu/~murray/synbio/wiki/index.php?title=Main_Page&direction=prev&oldid=52 AND gates used to build an adder (oligo technology, Winfree lab) http://www.cds.caltech.edu/%7Emurray/synbio/wiki/images/5/55/Chen-surf05.pdf
Massive models: http://www.cds.caltech.edu/%7Emurray/synbio/wiki/images/4/44/Ho-surf05.pdf
Cambridge http://www.ccbi.cam.ac.uk/iGEM2005/index.php/Main_Page Used sender/pulse-generator from Princeton to do something? AHL signal and aTc activated promoter Important paper in PNAS where this is shown to work: http://www.princeton.edu/~rweiss/papers/basu-pulse-2004.pdf
Harvard http://bio.freelogy.org/wiki/IGEM_2005 Bacterial wire propogates signal of AHL
MIT 2005 The first way we might build such a system involves the direct communication of an antigen, which can be just about anything, with the cell; this is accomplished by attaching an antibody to the cell in such a way that the binding of an antigen to the antibody initiates a signalling cascade that terminates in PoPs. The main benefit of such a system is that it can stand alone, and is thus a viable solution to problems such as "how do we deploy our biosensor into a lake where it can respond to toxin levels?" The main issue to be dealt with is that this system is in some ways less modular; of course, anyone could just follow our steps and hook up their scFv sequence of choice. http://openwetware.org/wiki/IGEM:MIT/2005/Direct_communication_of_antigen_and_receiver
UC Berkley 2005 http://parts2.mit.edu/wiki/index.php/UC_Berkeley_2005
Conjugation is a process through which cells can exchange genetic material on plasmids. Conjugal plasmids (in our case incF and incP plasmids) carry the machinery necessary to transfer themselves in the form of mating pair formation (mpf) and DNA transfer (dtr) genes. Conjugation is under the control of the TraJ regulatory protein, which when expressed induces a cascade that results in the formation of a pore by mpf genes and then subsequent nicking, rolling circle replication and transfer of one strand of the plasmid by the relaxosome complex and other dtr proteins. The relaxosome nicks the plasmid at the OriT region and then covalently attaches one of its subunits to the 5' end of the plasmid DNA, and by doing so it is able to drag the plasmid across the pore formed by the mpf machinery by means of a coupling protein. Upon reaching its destination, the single strand of plasmid DNA is recircularized and a complement strand is synthesized by transferred primases.
Non-mobile synthetic F plasmid: Begins the conjugation signal, which it sends to plasmid B. Also contains the CFP tag which identifies the host cell as "F-type", and always produces mRNA 'key 2' which unlocks RNA lock 2
-1. -B - Non-mobile almost-wild F plasmid: Contains all F-plasmid genes EXCEPT OriTf, TraJf. Plasmid receives and propagates the conjugation signal from TraJf in plasmid 1-A and sends the signal to OriTf in 1-C 1-C - Mobile F plasmid: Contains the OriTf site which receives signal from plasmid 1-B. This plasmid then leaves the host cell and enters the conjugating recipient cell. Holds encrypted message (produce cI --> turn on GFP to signify "message 1 received") secured by RNA lock 1.
2-A Non-mobile synthetic R plasmid: Always produces mRNA 'key1'. Thus when it receives 'lock1' (sent by mobile plasmid 1-C) it can open the latter and produce cI, which will activate plasmid 1-C (turn on GFP, "message 1 received") and simultaneously activate TraJr (start R conjugation cascade)
-1. 2-B Non-mobile almost-wild R plasmid: Just like 1-B, contains all of the wild type R-plasmid EXCEPT OriTr and TraJr. Propagates TraJr signal from 2-A and sends it to OriTr 2-C Mobile R plasmid: Contains the OriTr site, which receives signal from plasmid 2-B. This plasmid then leaves the host cell and submits its message back into cell #1
Penn State
http://parts2.mit.edu/wiki/index.php?title=Penn_StateProjectDes
”The idea for our project grew out of one for a "bacterial maze," in which bacteria would use logic to make their way through a microfabricated labrynth. This seemed slightly too difficult, so we linearized the the concept and added transfer of a signal; the idea was then dubbed a "bacterial relay race." As in a conventional relay race, the signal is to "go," or induce motility of a latter stage participant. This is accomplished by passing a baton. In our case, the participants are E. coli, and the baton is a quorum sensing molecule, 3OC6HSL (we have another strategy that utilizes conjugation rather than quorum sensing to mediate the signal). In addition to passing the signal, though, the first participant must stop. We explored this option, but settled instead on terminating the first participant. In our design we really do kill the messanger.”
Arizona
“Water Color”
http://parts.mit.edu/wiki/index.php/University_of_Arizona_2006
Project Details “The current name of our project is "Water Color." It is a system that selectively expresses one of three florescence proteins. Each of the three florescence proteins will be expressed in the presence of a unique inducer. Each florescent protein will be controlled by a unique repressed promoter. Thus we will have the expression of three flourescent proteins activated by the presence of there respective inducers. The idea of our project is to have a media with these cells on it so that each cell will be individually activated to shown a certain "color" (in actuallity, express one florescent protein, which may or may not look unique). Thus the media is able to dispaly an image. The spacial resolution with determine how much it will look like an image. A further idea, to be implemented later (time permitting), is to have the ability to "erase" the image. This would be accomplished by repressing all three promoters. Currently, there are no plans to implement this.”
Flowchart of Parts: http://parts.mit.edu/wiki/index.php/University_of_Arizona_2006/Parts_Schedule
Harvard
http://bio.freelogy.org/wiki/IGEM_2005
UC Berkley 2005 http://parts2.mit.edu/wiki/index.php/UC_Berkeley_2005
Conjugation is a process through which cells can exchange genetic material on plasmids. Conjugal plasmids (in our case incF and incP plasmids) carry the machinery necessary to transfer themselves in the form of mating pair formation (mpf) and DNA transfer (dtr) genes. Conjugation is under the control of the TraJ regulatory protein, which when expressed induces a cascade that results in the formation of a pore by mpf genes and then subsequent nicking, rolling circle replication and transfer of one strand of the plasmid by the relaxosome complex and other dtr proteins. The relaxosome nicks the plasmid at the OriT region and then covalently attaches one of its subunits to the 5' end of the plasmid DNA, and by doing so it is able to drag the plasmid across the pore formed by the mpf machinery by means of a coupling protein. Upon reaching its destination, the single strand of plasmid DNA is recircularized and a complement strand is synthesized by transferred primases.
Non-mobile synthetic F plasmid: Begins the conjugation signal, which it sends to plasmid B. Also contains the CFP tag which identifies the host cell as "F-type", and always produces mRNA 'key 2' which unlocks RNA lock 2
-1. -B - Non-mobile almost-wild F plasmid: Contains all F-plasmid genes EXCEPT OriTf, TraJf. Plasmid receives and propagates the conjugation signal from TraJf in plasmid 1-A and sends the signal to OriTf in 1-C 1-C - Mobile F plasmid: Contains the OriTf site which receives signal from plasmid 1-B. This plasmid then leaves the host cell and enters the conjugating recipient cell. Holds encrypted message (produce cI --> turn on GFP to signify "message 1 received") secured by RNA lock 1.
2-A Non-mobile synthetic R plasmid: Always produces mRNA 'key1'. Thus when it receives 'lock1' (sent by mobile plasmid 1-C) it can open the latter and produce cI, which will activate plasmid 1-C (turn on GFP, "message 1 received") and simultaneously activate TraJr (start R conjugation cascade)
-1. 2-B Non-mobile almost-wild R plasmid: Just like 1-B, contains all of the wild type R-plasmid EXCEPT OriTr and TraJr. Propagates TraJr signal from 2-A and sends it to OriTr 2-C Mobile R plasmid: Contains the OriTr site, which receives signal from plasmid 2-B. This plasmid then leaves the host cell and submits its message back into cell #1